Существенными эволюционными прогрессивными приобретениями птиц являются. Домашняя работа птицы тест. III группа. Кровеносная система, обмен веществ
Эволюция органического мира - длительный и сложный процесс, осуществляющийся на разных уровнях организации живой материи и протекающий в разных направлениях. Развитие живой природы происходило от низших форм, имеющих относительно простое строение, к все более усложняющимся формам. Одновременно внутри отдельных групп организмов развивались специальные приспособления (адаптации), позволяющие им существовать в конкретных местообитаниях. Например, у многих водных животных появляются перепонки между пальцами, облегчающие плавание (тритоны, лягушки, утки, гуси, утконос и др.).
Анализируя историческое развитие органического мира и многочисленные конкретные адаптации, крупнейшие российские эволюционисты А. Н. Северцов и И. И. Шмальгаузен выделили три главных направления эволюции: ароморфоз, идеоадаптации и дегенерация.
Ароморфозом (или арогенезом) называют крупные эволюционные изменения, ведущие к общему усложнению строения и функций организмов и позволяющие последним занимать принципиально новые местообитания или существенным образом повышающие конкурентную способность организмов в существующих местообитаниях. Ароморфозы позволяют переходить в новые среды обитания (то есть выходить в новые адаптационные зоны). Поэтому ароморфозы - сравнительно редкие явления в живом мире и носят принципиальный характер, оказывая большое влияние на дальнейшую эволюцию организмов.
Адаптационным уровнем или адаптивной зоной называется определенный тип местообитаний с характерными для него экологическими условиями или комплекс определенных адаптаций, свойственный той или иной группе организмов (общие условия жизни или сходные способы усвоения каких-то жизненно важных ресурсов). Например, адаптивной зоной птиц является освоение воздушного пространства, обеспечившее им защиту от многих хищников, новые способы охоты за летающими насекомыми (где у них нет конкурентов), быстрое передвижение в пространстве, возможности преодоления крупных препятствий, недоступных для других животных (реки, моря, горы и т. п.), способность к дальним миграциям (перелеты) и т. п. Поэтому полет является крупным эволюционным приобретением (ароморфозом).
Наиболее яркими примерами ароморфозов могут служить многоклеточность и появление полового способа размножения. Многоклеточность способствовала появлению и специализации тканей, привела к усложнению морфологии и анатомии многих групп организмов, как растений, так и животных. Половое размножение существенно расширило адаптационные способности организмов (комбинативная изменчивость).
Ароморфозы обеспечили животным более эффективные способы питания и повысили эффективность обмена веществ - например, появление челюстей у животных позволило перейти от пассивного питания к активному; освобождение пищеварительного канала от кожно-мускульного мешка и появление в нем выводного отверстия принципиально улучшило эффективность усвоения пищи за счет специализации разных его отделов (появление желудка, отделов кишки, пищеварительных желез, быстрый вывод ненужных продуктов). Это существенно повысило возможности выживания организмов даже в местах с малым содержанием питательных ресурсов.
Крупнейшим ароморфозом в эволюции животных стала теплокровность, резко активировавшая интенсивность и эффективность обмена веществ у организмов и повысившая их выживаемость в местообитаниях с низкой или резко меняющейся температурой.
В качестве примеров ароморфозов в животном мире можно вспомнить также образование внутренней полости организмов (первичной и вторичной), появление скелета (внутреннего или внешнего), развитие нервной системы и особенно усложнение строения и функций головного мозга (появление сложных рефлексов, обучаемости, мышления, второй сигнальной системы у человека и т. д.) и многие другие примеры.
У растений крупными ароморфозами являются: появление проводящей системы, связавшей разные части растения в единое целое; образование побега - жизненно важного органа, обеспечившего растениям все стороны жизнедеятельности и воспроизводства; формирование семени - органа воспроизводства, возникающего половым путем, развитие и созревание которого обеспечивается ресурсами всего материнского организма (дерева, кустарника или иной жизненной формы растений) и который имеет хорошо защищенный тканями семени зародыш (голосеменные и покрытосеменные растения); появление цветка, повысившего эффективность опыления, уменьшившего зависимость опыления и оплодотворения от и обеспечившего защиту яйцеклетки.
У бактерий ароморфозом можно считать появление автотрофного способа питания (фототрофного и литотрофного или хемосинтетического), позволившего им занять новую адаптационную зону - местообитания, полностью лишенные органических источников питания или имеющие дефицит таковых. У бактерий и грибов к ароморфозам можно отнести способность образовывать те или иные биологически активные соединения (антибиотики, токсины, ростовые вещества и т. п.), существенно повышающие их конкурентную способность.
Арогенез может происходить и на межвидовом (или биоценотическом) уровне при взаимодействии организмов разного систематического положения. Например, появление перекрестного опыления и привлечение для этого насекомых и птиц можно рассматривать как ароморфоз. Крупными биоценотическими ароморфозами являются: формирование микориз (симбиоз грибов и корней растений) и лишайников (объединение грибов и водорослей). Эти типы ассоциаций позволили симбионтам обитать в таких местах, где порознь они никогда бы не поселились (на бедных почвах, на скалах и т. п.). Особенно знаменателен союз грибов и водорослей, приведший к появлению новой симбиотической формы жизни - лишайников, которые морфологически очень похожи на единый организм, напоминающий растения. Крупнейшим ароморфозом этого типа является эукариотическая клетка, состоящая из разных организмов (прокариот), полностью потерявших свою индивидуальность и превратившихся в органеллы. Эукариотическая клетка обладает более активным и экономичным обменом веществ по сравнению с прокариотической и обеспечила появление и эволюцию царств грибов, растений и животных.
Ароморфозы - это крупные события в эволюции органического мира, и они сохраняются в популяциях и в дальнейшем развитии приводят к появлению новых крупных групп организмов и таксонов высокого ранга - порядков (отрядов), классов, типов (отделов).
Предполагается, что ароморфоз наиболее вероятен у исходно примитивных или мало специализированных форм организмов, так как они легче переносят изменения окружающей среды и им проще освоиться в новых средах обитания. Специализированные формы, приспособленные к определенным, часто достаточно узко ограниченным условиям жизни, обычно погибают при резком изменении таких условий. Именно поэтому в природе, наряду с высокоорганизованными и специализированными формами жизни, сосуществует большое число сравнительно примитивных организмов (бактерий, грибов, беспозвоночных и прочих), прекрасно адаптировавшихся в новых условиях и очень устойчивых. Такова логика эволюционного процесса.
Общая дегенерация, или катагенез
Это конкретные адаптации к определенным специфическим условиям обитания, образующимся в пределах одной и той же адаптационной зоны. Идиоадаптации проявляются как при арогенезе, так и при дегенерации. Это частные приспособления, не меняющие существенно уровня организации организмов, достигнутого в процессе эволюции, но заметно облегчающие их выживание именно в данных местообитаниях.
Например, если цветок мы можем рассматривать как крупнейший ароморфоз в эволюции растительного мира, то формы и размеры цветка определяются уже теми реальными условиями, в которых существуют те или иные виды растений, или их систематическим положением.
То же касается, например, птиц. Крыло - это ароморфоз. Форма крыльев, способы полета (парящий, маховотолчковый) - серия идиоадаптаций, не меняющая принципиально морфологической или анатомической организации птиц. К идиоадаптациям можно отнести покровительственную окраску, широко распространенную в мире животных. Поэтому идиоадаптации нередко рассматривают как признаки низших таксономических категорий - подвидов, видов, реже родов или семейств.
Соотношение различных направлений в эволюции
Эволюционный процесс происходит непрерывно, и основные его направления могут меняться со временем.
Ароморфозы или общая дегенерация, как редкие процессы в эволюции, приводят к повышению или понижению морфолого-физиологической организации организмов и занятию ими более высокой или низкой адаптивной зоны. Внутри этих адаптивных зон начинают активно развиваться частные приспособления (идиоадаптации), обеспечивающие более тонкое приспособление организмов к конкретным местообитаниям. Например, появление большой группы микоризообразующих грибов позволяет им занять новую адаптационную зону, связанную с большой группой новых для грибов и растений местообитаний. Это биоценотический ароморфоз, сопровождающийся далее серией частных приспособлений (идиоадаптаций) - расселением разных видов грибов по разным растениям-хозяевам (подберезовики, подосиновики, боровики и т. д.).
В процессе эволюции биологический прогресс может сменяться регрессом, ароморфозы - общей дегенерацией, и все это сопровождается новыми идиоадаптациями. Каждый ароморфоз и каждая дегенерация вызывают расселение организмов по новым средам обитания, реализуемое через идиоадаптации. Таково соотношение этих направлений эволюционного процесса. На основе данных эволюционных преобразований организмы занимают новые экологические ниши и заселяют новые местообитания, то есть происходит их активная адаптивная радиация. Например, выход позвоночных животных на сушу (ароморфоз) вызвал их адаптивную радиацию и привел к формированию многих таксономических и экологических групп (хищники, травоядные, грызуны, насекомоядные и т. п.) и новых таксонов (амфибии, рептилии, птицы, млекопитающие).
Общая характеристика направлений эволюции по изменению уровня организации и характеру процветания вида.
Конвергенция и дивергенция
Анализ механизма видообразования показывает, что результатом этого процесса является появление одного или нескольких (двух, трех и более) близких видов.
Рассматривая эволюцию в целом, можно увидеть, что результатом ее является все многообразие организмов, живущих на Земле. Поэтому, основываясь на результатах эволюционного процесса, можно выделить два вида эволюции - микроэволюцию и макроэволюцию.
Микроэволюция - совокупность процессов видообразования, при которых из одного вида возникают новые (один или несколько) видов организмов.
Микроэволюция представляет собой как бы «элементарный акт эволюции», сопровождающийся появлением небольшого количества видов из одного исходного вида.
Примером микроэволюционных процессов является возникновение двух рас ночной бабочки Березовая пяденица, разных видов вьюрков на Галапагосских островах, прибрежных видов чаек на побережье Северного Ледовитого океана (от Норвегии до Аляски) и т. д.
Выведение породы «белая украинская свинья» может служить примером микроэволюции, реализуемой человеком.
Таким образом, результатом микроэволюции является появление новых видов из исходного вида, которое осуществляется за счет дивергенции.
Дивергенция - это процесс расхождения признаков, в результате которого появляются новые виды или возникшие в процессе эволюции виды отличаются друг от друга различными признаками за счет приспособления этих видов к разным условиям существования.
Макроэволюция - совокупность всех эволюционных процессов, в результате которых возникло все многообразие органического мира; эти процессы идут не только на уровне вида, но и на уровне рода, семейства, класса и т. д.
Результатом макроэволюции является все многообразие современного органического мира, которое возникло как за счет дивергенции, так и за счет конвергенции (схождения признаков).
Виды, возникшие из разных групп организмов (например, классов), могут быть конвергентными, т. е. наряду с определенными различиями они имеют общие признаки, связанные с приспособленностью к одной среде обитания. Примером конвергентных видов являются акула, кит и ихтиозавр (ископаемое пресмыкающееся). Эти виды имеют рыбообразную форму, плавники, так как приспособлены к водной среде. Другим примером конвергентных организмов являются бабочки, птицы и летучие мыши, так как они имеют крылья и приспособлены к воздушно-наземному образу жизни.
Следовательно, при протекании макроэволюции возможны и дивергенция, и конвергенция.
В течение длительного исторического развития макроэволюция приводила к резкому изменению органического мира в целом. Так, современный органический мир значительно отличается от такового для протерозойской или мезозойской эр.
Пути и направления эволюции
Как было отмечено выше, эволюция осуществляется двумя способами - дивергентным и конвергентным, и в результате этих процессов возникают различные виды как по уровню их организации, так и по характеру приспособления к средам обитания. Поэтому различают три пути эволюции по характеру изменения уровня организации возникающих организмов: идиоадаптация, ароморфоз и дегенерация.
1. Ароморфоз (арогенез) - путь эволюции, при котором уровень организации организмов по сравнению с исходными формами повышается.
К ароморфозам относятся: возникновение фотосинтезирующих организмов из гетеротрофов; появление многоклеточных организмов из одноклеточных; возникновение псилофитов из водорослей; появление покрытосеменных с наличием двойного оплодотворения и новых оболочек у семени из голосеменных; возникновение организмов, способных вскармливать своих детенышей молоком, и т. д.
2. Идиоадаптация (аллогенез) - путь эволюции, при котором появляются новые виды, по уровню организации не отличающиеся от исходных видов.
Возникшие при идиоадаптациях виды отличаются от исходных признаками, позволяющими нормально существовать в различных условиях обитания. К идиоадаптации можно отнести появление разных видов вьюрков на Галапагосских островах, различных грызунов, живущих в разных условиях (зайцы, суслики, мышевидные грызуны), и другие примеры.
3. Дегенерация (катагенез) - путь эволюции, при котором общий уровень вновь возникших организмов понижается.
В некоторых источниках пути эволюции называют направлениями. В этом случае необходимо указывать: направления эволюции по характеру изменения уровня организации, так как существуют направления эволюции по характеру процветания. По данному признаку различают два направления - биологический прогресс и биологический регресс.
Биологический прогресс - это такое направление эволюции, при котором увеличивается число популяций, подвидов и расширяется ареал (территория обитания), при этом данная группа организмов находится в состоянии постоянного видообразования.
В настоящее время в состоянии биологического прогресса находятся млекопитающие, членистоногие (из животных), покрытосеменные (из растений). Биологический прогресс не означает повышения уровня организации организмов, но и не исключает его.
Биологический регресс - направление эволюции, при котором уменьшаются ареал и численность организмов, замедляются темпы видообразования (уменьшается число популяций, подвидов, видов).
В настоящее время в состоянии биологического регресса находятся пресмыкающиеся, земноводные (из животных), папоротникообразные (из растений). При этом большое влияние на состояние прогресса или регресса организмов оказывает деятельность человека. Так, многие виды животных вымерли из-за воздействия человека (например, тюлень Стеллерова корова, туры и т. д.).
Приспособленность организмов к условиям среды, ее виды и относительность
Первое научно обоснованное определение вида дал Ч. Дарвин. В настоящее время это понятие уточнено с позиций всех современных теорий, в том числе и с генетической позиции. В современной интерпретации формулировка понятия «вид» такова:
Вид - это совокупность всех особей, обладающих одинаковыми наследственными морфологическими и физиологическими признаками, способных свободно скрещиваться и давать нормальное плодовитое потомство, имеющих одинаковый геном, одинаковое происхождение, занимающих определенный жизненный ареал и приспособленных к условиям существования в нем.
Критерии вида и его экологическая характеристика будут рассмотрены далее. В данном подразделе приведем механизм видообразования.
В пределах популяций у различных особей этих популяций за счет мутационной (наследственной) изменчивости возникают различные признаки, поэтому все особи данной популяции имеют определенные отличия друг от друга.
Признаки, появляющиеся у отдельных особей, могут быть либо полезными, либо вредными для этого организма в условиях данной среды обитания. В процессе жизнедеятельности, как правило, выживают те особи, которые более приспособлены к данной среде обитания. У особей разных популяций эти признаки будут различны особенно в случае, когда условия сред их обитания будут сильно отличаться.
С течением времени признаки, отличающие особей одной популяции от другой, накапливаются, и различия между ними становятся все более и более значительными. В результате этих процессов из одного исходного вида возникает несколько подвидов (их число таково, сколько было популяций вида, живущих в разных условиях среды, - 2, 3 и т. д.).
В случае, если разные популяции, находящиеся в разных условиях существования, достаточно сильно изолированы друг от друга, то смешивания признаков из-за гибридизации особей не происходит. Различия между особями разных популяций становятся столь значительными, что можно констатировать возникновение новых видов (их особи уже не скрещиваются и не дают полноценного плодовитого потомства).
В процессе видообразования возникают новые виды, которые оказываются хорошо приспособленными к условиям своего существования, что всегда удивляло и восхищало человека, а религиозных людей заставляло восхищаться «мудростью творца». Рассмотрим сущность явления приспособленности, а также относительность приспособленности.
Приспособленностью называют определенные признаки организмов, позволяющие ему выжить в данных конкретных условиях среды обитания.
Ярким примером приспособленности является белая окраска зайца-беляка в зимний период. Эта окраска делает его незаметным на фоне белого снежного покрова.
В процессе эволюции у многих организмов выработались признаки, благодаря которым они очень хорошо приспособились к среде своего обитания. Эволюционная теория вскрыла причину и механизм возникновения приспособленности организма к условиям среды своего обитания, показало материалистическую сущность этого процесса.
Причиной появления приспособлений к условиям среды обитания является наследственная изменчивость, возникающая под влиянием условий окружающей среды.
Возникшие мутации в случае их полезности закрепляются в потомстве за счет лучшего выживания особей, обладающих этими признаками.
Классический пример возникновения приспособленности у организмов к среде обитания был показан в трудах Ч. Дарвина.
В Англии обитает ночная бабочка Березовая пяденица, имеющая светло-желтую окраску. На фоне светлого ствола березы эти бабочки незаметны, поэтому большинство из них сохраняются, поскольку незаметны для птиц.
Если березы растут в районе предприятия, выделяющего копоть, то их стволы темнеют. На их фоне светлоокрашенные бабочки становятся заметными, поэтому легко поедаются птицами. В процессе длительного временного существования вида этих бабочек, за счет мутаций появились формы с темной окраской. Темноокрашенные формы в новых условиях лучше выживали, чем светлоокрашенные. Так, в Англии возникли два подвида бабочек-пядениц (светло- и темноокрашенные формы).
Реконструкция производства и улучшение технологии с учетом требований привели к тому, что предприятия перестали выделять копоть и изменять окраску стволов берез. Это привело к тому, что темноокрашенные формы для новых условий стали не приспособлены, и признак, приобретенный ими, стал не только не полезным, а даже вредным. На этом основании можно сделать вывод о том, что приспособленность организмов относительна: сильное, даже кратковременное, изменение условий среды может организм, приспособленный к среде обитания, превратить в неприспособленный: например, заяц-беляк при слишком раннем сходе снежного покрова будет сильнее заметен на фоне темного поля, чем если бы он был окрашен в «летний» (серый) цвет.
Существует несколько разновидностей приспособленности организмов. Рассмотрим некоторые из них.
1. Покровительственная окраска - окрас, позволяющий организму быть незаметным на фоне окружающей среды.
Примеры: зеленая окраска тли на фоне зеленых листьев капусты; темная окраска спины рыбы на темном фоне при взгляде сверху и светлая окраска брюха на светлом фоне при взгляде снизу; рыбы, живущие в зарослях водной растительности, имеют полосатую окраску (щука) и т.д.
2. Мимикрия и маскировка.
Мимикрия состоит в том, что организм по форме похож на другой организм. Примером мимикрии является муха осовидка, форма ее тела напоминает осу и этим предостерегает об опасности, которой нет, так как эта муха не имеет жала.
Маскировка состоит в том, что организм приобретает форму какого-то предмета окружающей среды и становится незаметным.
Примером могут служить палочники - насекомые, по форме напоминающие обломки стеблей растений; есть насекомые, имеющие листообразную форму и т. д.
3. Предупреждающая окраска - яркая окраска, предупреждающая об опасности. Примеры: окраска ядовитых божьих коровок, пчел, ос, шмелей и т.д.
4. Особые приспособления растений для реализации процессов опыления. Ветроопыляемые растения имеют длинные, свисающие тычинки, удлиненные, торчащие в разные стороны рыльца пестиков с приспособлениями для улавливания пыльцы и другие формы. Насекомоопыляемые растения имеют соцветия, яркую краску и экзотические формы цветка для привлечения определенного вида насекомого, с помощью которого реализуется опыление.
5. Особые формы поведения животных - угрожающие позы порой безобидных, а иногда и опасных рептилий, зарывание страусом головы в песок и т. д.
Подводя итог, можно отметить, что благодаря накоплению различий, возникающих за счет мутаций, возможно образование новых видов, приспособленных к среде своего обитания, но эта приспособленность относительна, так как изменение условий приводит к утере приспособленности организма к данной среде.
Результаты недавно проведённого, всестороннего кладистического исследования эволюции птиц под руководством Марьянской и группы учёных лишь подлили масла в огонь. Основываясь на анализе многих важных образцов, находящихся в Польше, Монголии и России, и на основе 195 скелетных признаков теропод и ранних птиц, было проведено доскональное исследование взаимосвязи между тероподами и ранними птицами. Другое недавно опубликованное исследование под руководством Жоу и Жанга, сделанное на основе 201 скелетных признаков, дополняет первое. Оно предусматривает отслеживание самой эволюции птиц, измеряя предполагаемый эволюционный континуум, который существует между dromaeosaurids как корневым видом, с одной стороны, и современными птицами как наивысшей группой с другой стороны. (Обратите внимание, что корневой вид сразу же оказывается за пределами предполагаемой эволюционной прогрессии, и используется в качестве точки отсчета для “примитивного” состояния, тогда как наивысшая группа состоит из наиболее выведенных членов предполагаемой эволюционной прогрессии).
Типичные эволюционные утверждения, поддерживающие существование “переходных форм”, основаны на избирательных данных - которые обычно заключаются в постепенном появлении внешних признаков, или же в развитии только одного или нескольких признаков. Наоборот, анализ данных, используемых для построения кладограммы, предусматривает одновременное сравнение всех значимых признаков, которые отличаются между “примитивными” и “произошедшими” существами. В данной работе проводится анализ новых исследований и используется та же самая методология, которая использовалась в ранних исследованиях предполагаемых эволюционных переходов: от предков человека - к появлению человека, от млекоподобных рептилий – к млекопитающим, от рыб - к амфибиям, и от млекопитающих, обитающих на суше - к китам.
Метод анализа
Несмотря на то, что и выбор характеристик, и некоторые аспекты их оценки содержат определённый элемент субъективности, набор данных используемых для кладистических анализов предусматривает относительно объективный, частично количественный анализ предполагаемых эволюционных изменений. Как обычно, в кладистических анализах подавляющее большинство анатомических признаков подсчитываются как противоположности по критерию «присутствия» или «отсутствия» (1 или 0) в каждом организме. Лишь некоторые признаки засчитываются в соответствии с возрастающим появлением признака (0, 1, 2, 3, и т.д.).
Таблица 1. Предположительный ход эволюции от теропод до Археоптерикса и обратно к существам, обитающим на суше.
В кладистических исследования, приведенных здесь, организмы выстроены как “указатели”, которые ведут к последовательному появлению птичьих признаков, но без необходимых взаимоотношений между предками и потомками. Клады и их соответствующие составные элементы приводятся в Таблицах 1–4. В Таблице 1 четыре группы теропод служат одной точкой отсчёта (корневой группой), и современные птицы (не показанные ни в цитируемом исследовании, ни в Таблице 1) другой точкой отсчёта. Для того чтобы избежать рассуждений по кругу относительно какого-либо предположения о существовании общего эволюционного процесса, который группирует ряд признаков (и затем сказать, что получившаяся кладограмма эволюционного дерева подтверждает эту предполагаемую эволюционную последовательность), каждый из 195 признаков данного исследования был в равной степени оценен и подсчитан, чтобы быть независимым друг от друга. Обратите внимание, что Клад Avialae (Таблица 1) соответствует общепринятому названию “птицы”. Порядок кладов, показанный в Таблице 1, изменяется в обратном направлении от начальных данных, так что предполагаемый ход эволюции развивается вверх, синхронно с ходом эволюции, показанном в Таблицах 2–4. Существующие в настоящее время птицы, к которым относятся утки и куры, присоединены к Классу Птиц в Таблице 2 и 4. Эти пять птиц составляют наивысшую группу.

Таблица 2. Линия эволюции птиц с акцентом на летающих птицах.
Каждый перечисленный таксон в Таблице 1 является примитивной родственной группой всего скопления таксонов, перечисленных выше его. В отличие от этого, ход эволюции летающих птиц (Таблица 2) имеет менее прямую, гнездовую ветвящуюся последовательность. Только шесть основных таксонов каждый составляет примитивную родственную группу из всех объединенных таксонов, расположенных выше них. Группа Gobipteryx-Cathayornis (являющаяся сама по себе сложно разветвлённой внутри) является родственной группе Patagopteryx - современные птицы (сама сложно разветвлённая внутри), и считается, что группа Confuciusornis предположительно является примитивной родственной группой обеих объединенных групп. В Таблице 3 связь простой родственной группы с каждым основным таксоном по отношению к её последователям (идентична только нижней части Таблицы 1) устраняет необходимость описания разветвлённых структур. Однако сложная разветвлённая взаимосвязь, которая свойственна завершающим стадиям эволюции птиц (Таблица 4) вынуждает добавление этих структур, как это и было сделано в Таблице 2.

Таблица 3. Появление ранних птиц: сокращённый вариант Таблицы 2.
Основная проблема использования второго набора данных заключается в очень большой порции информации, которой не хватает. Для того чтобы уменьшить вероятность отклонений, представленных в колонке Птичьи Признаки, при оценивании имеющихся данных были использованы два разных подхода. Для того чтобы сохранить информацию, которая относится ко всему ходу эволюции летающих птиц, была сохранена полная последовательность от Dromaeosauridae до современных птиц, несмотря на сильную потерю данных (оказалось, что лишь 37 признаков из 201 используемы, из которых 21 был невозвратные), как представлено в Таблице 2. В результате значительной потери данных, а также из-за того, что клад Gobipteryx-Cathayornis составляет большую часть этой потери, этот клад был исключен из дальнейшего рассмотрения. Оставшаяся информация была разделена на Раннюю (Таблица 3) и Позднюю (Таблица 4) эволюции летающих птиц. Это сильно уменьшило потерю данных, так как каждый набор данных теперь мог быть упорядочен только в соответствии с некоторыми отсутствующими точками данных, которые встречаются во время коротких промежутков этого ряда значений. В Таблице 3 и 4, из исходных 201 очков данных теперь оценивают для анализа 131.
Анализ взаимосвязей между тероподами и птицами
На всём протяжении последовательности тероподы-птицы (Таблица 1), прослеживается почти монотонное последовательное появление птичьих признаков. Однако существует видимое возвратное изменение в Птичьих Признаках у теропод, которое непосредственно предшествует первой известной общепризнанной птице, Археоптериксу . Кроме того, более тщательный анализ данных указывает на то, что видимая плавность общей последовательности на самом деле обманчива. Во-первых, как всегда, процесс упорядочивания сам по себе помогает эволюционистам. Более того, добавление возвратных признаков сглаживает общую последовательность. Поразительно, 140 из всех 195 признаков изменяются обратно по меньшей мере один раз, и если включить четыре корневые тероподовые группы, то это число увеличивается до 145. Кроме того, из 140 возвратных признаков, которые прослеживаются внутри последовательности, 64 изменяются в обратном направлении, по меньшей мере, дважды. Следовательно, большинство основных птичьих признаков не прогрессируют в сторону птичьего состояния! Наоборот, перед нами смешанная коллекция мозаики, которая состоит из птиц и рептилий.
Теперь рассмотрим только прогрессивные признаки. Эта последовательность характеризуется резкими скачками в предполагаемом приобретении птичьих признаков. Обратите внимание, например, на удваивание Птичьих Признаков почти в два раза с соответствующим появлением Eumaniraptora и “C” кладов. Также существует большой разрыв между Dromaeosauridae и Troodontidae . Два клада, которые непосредственно предшествуют Археоптериксу , по иронии судьбы, в морфологическом строении скелета более птицеподобные (в отношении современных птиц), чем сам Археоптерикс !
Эволюция в неверном направлении
Часть последовательности после Археоптерикса (Таблица 1), предложенная только несколькими более ранними исследователями, вновь находит свою поддержку. Только представьте всю ироничность этой ситуации: определённые нелетающие “тероподы” (oviraptorids ), включая известного “пернатого теропода” Каудиптерикса, оказались более птицеподобными, чем летающий Археоптерикс :
“Некоторые особенности черепа, которые наблюдаются у oviraptorids (нет сведений, что у других oviraptorids также присутствует такая особенность черепа) поддерживают нашу гипотезу относительно птичьего статуса Oviraptorosauria …. Эта группа признаков отсутствует у нелетающих тероподов, но присутствует у развитых птиц …. Несмотря на эти схожести с летающими птицами, oviraptorosaurs не имеют каких-либо приспособлений к полёту в своей заднечерепной части [слова курсивом были добавлены].”

Таблица 4. Наиболее современные стадии (включая окончательную группу) эволюции птиц: сокращенный вариант Таблицы 2.
(Обратите внимание, что прилагательное avialan относится к кладу avialidae, который, как было указано ранее, включает всех вымерших и современных птиц). Смесь птичьих и не птичьих признаков, встречающихся у oviraptorosaurs , можно отстаивать только путём отдельной эволюционной линии для oviraptorosaur , отходящей от основной линии происхождения птиц, которая начинается с Археоптерикса , с последующими многочисленными эволюционными возвратными признаками среди этого предполагаемого “бокового ответвления” эволюции:
«Если считать такую связь правдоподобной, то в таком случае oviraptorosaurs были не способны к полёту. Следовательно, некоторые заднечерепные признаки oviraptorosaurs в ходе анализа признаются как reversals возвратные изменения. Примерами таких возвратных изменений являются… (перечисляются некоторые признаки, которые предположительно указывают на то, что oviraptorosaurs “перешли” обратно в нелетающее состояние). Эти возвратные изменения, очевидно, сопровождали изменение из летающего способа жизни к обитанию на суше»
Вряд ли кто-то удивится, узнав, что этот эволюционный рассказ совсем не подтверждается летописью окаменелостей:
«В данный момент трудно предложить сценарий, который бы описывал последовательные ступени эволюции от летающих птиц к нелетающим oviraptorosaurs. И, тем не менее, накопленные свидетельства признаков указывают на то, что такое радикальное изменение адаптации, из летающего в обитающего на земле, возможно, произошло впервые на раннем этапе эволюции птиц»
В результате эволюционисты, которые не способны предоставить необходимые доказательства, оказались в ещё более затруднительном положении. Им не только не хватает постепенного появления основных приспособлений для полёта, но теперь им также не хватает и постепенного исчезновения этих приспособлений в случае с “второстепенными нелетающими” oviraptorosaurs !
Тероподы и неудачный аргумент стратоморфических промежуточных звеньев
Некоторые эволюционисты настойчиво утверждают, что эволюция можно считать достоверной хотя бы только по той причине, что окаменелости с “промежуточными структурами” всегда можно обнаружить в соответствующей части геологической колонны. К сожалению, некоторые креационисты также поддались на эту уловку ложного рассуждения. Аргумент стратоморфического промежуточного звена можно было бы считать действительным, если бы: (1) временной стратиграфический промежуток в обсуждаемом вопросе содержал бы только одну группу потенциально рассматриваемых структурных промежуточных звеньев, а также, если бы (2) предполагаемые структурные промежуточные звенья появлялись бы только в соответствующем стратиграфическом промежутке, где они необходимы в соответствии с эволюционной теорией (т.е. соединить две другие группы в эволюционной последовательности).
Давайте рассмотрим млекоподобных рептилий. Они, вероятно, являются главным примером стратоморфических промежуточных звеньев. Но представьте, что бы случилось, если бы они никогда не существовали или никогда не были бы обнаружены. Тогда эволюционисты вслед за “Бульдогом Дарвина”, Томасом Гексли, вероятно, ссылались бы на древних амфибий как на потомственную группу млекопитающих. Какая-нибудь вымершая группа амфибий была бы описана как стратоморфические промежуточные звенья, которые соединяют не-млекопитающих и млекопитающих. Это нарушает первое условие. В данном исследовании тероподы вскрывают ошибочность стратоморфических промежуточных звеньев, так как прямо противоречит второму условию. Они являются замечательным примером организмов, которые в некоторой степени подходят на роль морфологических промежуточных звеньев в предполагаемой эволюционной последовательности, но которые появляются в неправильной части геологической колонны для того, чтобы быть переходными формами.
Существует ряд свидетельств, которые указывают на то, что тероподы, предшествующие птицам (Таблица 1), появляются слишком поздно в стандартной геологической колонне (Юрский период) для того, чтобы считаться предками птиц. Например, несмотря на то, что Протоптица Protoavis появилась предположительно десятки миллионов лет до Археоптерикса , имеет сходство с современными птицами в большей степени, чем с Археоптериксом . Недавнее обнаружение птицеподобных отпечатков также является веским доказательством того, что птицы появились в стандартной геологической колонне задолго до появления существ, перечисленных в Таблицах 1–4:
«Известная история птиц начинается в Эпоху Позднего Юрского периода (приблизительно 150 миллионов лет назад), именно тогда и датируют появление Археоптерикса … Здесь мы описываем множество хорошо сохранившихся отпечатков с явными птичьими признаками в красноцветных отложениях Аргентины, которые относятся к Позднему Триасовому периоду, по меньшей мере за 55 миллионов лет раньше первых известных скелетных остатков птиц»
Более ранние заявления относительно отпечатков, отнесенных к Позднему Триасовому периоду, были поставлены под вопрос, и в результате отпечатки приписали к нептицеподобным динозаврам. Однако упоминаемые авторы утверждают, что недавно обнаруженные отпечатки по структуре несравненно больше напоминают отпечатки птиц, чем ранние отпечатки.
В любом случае интересно то, что некоторые эволюционисты признают тот факт, что тероподы (включая перечисленных в Таблице 1) возможно даже и не подходят на роль непрямых предков птиц. Например, вот что говорит эволюционист Питер Добсон:
«Спешу заметить, что ни один из известных небольших теропод, включая Deinonychus, Dromaeosaurus, Velociraptor, Unenlagia, а также Sinosauropteryx, Protarcheaeopteryx, Caudipteryx сам по себе не имеет никакого отношения к происхождению птиц; все они являются окаменелостями, которые относятся к Меловому периоду … и являясь таковыми, могут в лучшем случае представлять только структурные стадии, через которые предположительно прошел предок птиц»
«Признаюсь, что я немного удивил сам себя. Когда какие-либо идеи становятся слишком популярными, а звук сирены новых иконоборческих идей становится слишком громким, Я упираюсь и начинаю смотреть на обратную сторону этой идеи. Я выступаю против кладистики и катастрофического вымирания динозавров; Я больше склонен к эндотермным динозаврам; Я скептически отношусь к идее того, что тераподы являются предками птиц»
Эволюция летающих птиц
Теперь обратим внимание на предполагаемую линию, которая достигает вершины в современных птицах как наивысшей группой. Если предположить, что Археоптерикс был первой птицей, тогда каким образом современные птицы предположительно эволюционировали из него? Если поместить его в контекст всей истории эволюции птиц (Таблица 2, “Все Признаки”), становится очевидным, что крупные разрывы последовательности в “Птичьем Признаке” как предшествовали Археоптериксу, так и следовали после него. Что касается только прогрессивных признаков, ни один не может быть отслежен в течении всего пути от предположительно потомственных dromaeosaurids полностью до современных птиц. Однако оставшиеся прогрессивные признаки в Таблице 2 указывают на второе крупное нарушение последовательности в эволюции ранних птиц - а именно между Confuciusornis и его примитивным родственником Sapeornis . С другой стороны существует большой разрыв (14.3 против 31.3) между Confuciusornis и наименее выведенными членами этой развитой родственной группы.
Тщательное рассмотрение предположительной ранней эволюции птиц (Таблица 3) под увеличительным стеклом лишь увеличивает и делает более явными разрывы в последовательностях. Относительно маленькая ступенька от dromaeosaurids к Археоптериксу , сглаженная в колонке “Все Признаки”, увеличивается в колонке “Прогрессивные Признаки”. “Птичий Признак” удваивается от dromaeosaurids к Археоптериксу , потом удваиваясь еще раз от Археоптерикса до Rahonavis . В колонке “Все Признаки” от Археоптерикса до Rahonavis существует пропасть (которая в четыре раза больше “Птичьего Признака”). От Sapeornis до Confuciusornis также существует большой скачок (который почти удваивает “Птичий Признак”), как в колонке “Все Признаки” так и в колонке “Прогрессивные Признаки” Таблицы 3. Если бы этого было не достаточно, то 21 из 131 полезных признаков, использованных в Таблице 3, изменялись возвратно в эволюционной последовательности, по меньшей мере, один раз.
От Археоптерикса до современных птиц
Конечно, данные, которые относятся к Археоптериксу (Таблица 1–3) не рассказывают всей истории. Интересно, что эволюционисты 19-го столетия, очевидно следуя здравому смыслу, признали тот факт, что Археоптерикс не считается настоящим видом, который заполняет пропуски большинства морфологических свойств, которые отделяют рептилий от птиц:
«Если взглянуть в прошлое, то кажется странным, что эволюционные теоретики долгое время находились под влиянием старинных представлений о гармонии. Исторически считалось, что многие окаменелости не являются частью потомственной линии, если они показывают смешивание ранних и поздних признаков, так как ожидалось, что промежуточные формы продемонстрируют идеальную промежуточность между более ранними и более поздними формами. Таким образом, такая окаменелость как Археоптерикс , показывающая смесь признаков рептилии и птиц, не могла быть размещена как переходная стадия между этими двумя классами, так как все эти отличительные признаки не являются переходными: считалось, что проходила путём постепенного и общего преобразования всего организма животного»
По мнению современных учёных-креационистов, среди всех известных видов не существует ни одного, ведущего к Археоптериксу, и у которого бы были структуры, напоминающие полукрыло и полуноги. Но несмотря на очевидное или действительное существование “теропод, имеющих оперенье”, предположительное эволюционное происхождение перьев остается проблематичным. В наше время считается, что происходит мозаично, и это предположительно оправдывается эволюционными изменениями в эволюционных путях имеющих отношение к эмбриональному развитию. Но, как мы отмечали ранее, эмбриональное развитие может совершенно противоречить общепринятым эволюционным схемам, в особенности учению “от теропод - к птицам”. Второй неизбежный факт: эволюционисты занизили свои стандарты относительно того, что можно считать доказательством. Не сумев найти окаменелых животных, которые бы показывали полную переходность между рептилиями и птицами, теперь они вынуждены склеивать вместе “серии” окаменелых животных, которые попросту показывают разнообразный набор признаков рептилий и птиц.
Самые недавние стадии эволюции птиц также наполнены разрывами последовательности и возвратными изменениями. Что касается последнего, 29 используемых признаков из 131, содержащихся в Таблице 4, изменяются в обратном направлении, по меньшей мере, один раз. Представьте, каким бы образом распределились Птичьи Признаки в Таблице 4, если бы они были плавно переходными. Показатели “Все Признаки” имели бы приблизительно следующие значения: от 50.5 до 62.8 до 75.1 и достигали бы верхнего предела 87.4 (современная птица имеет наименьшее значение в Птичьих Признаках в колонке “Все Признаки”). Соответствующие значения для Прогрессивных Признаков также начинались бы с 50.5, переходя к 66.7 и 83.4 до того, как достигали бы значения 100 (современная птица имеет наименьшее значение Птичьего Признака в колонке “Прогрессивные Признаки”). Реальность за Птичьими Признаками, относящимся к эволюции современных птиц (Таблица 4) совершенно другая. Между Patagopteryx и его развитой родственной группой современных птиц Apsaravis , существует сильный разрыв. Это нарушение последовательности появляется в обеих колонках общего обзора эволюции летающих птиц (Таблица 2), как и в обеих колонках детального обзора недавней эволюции птиц (Таблица 4).
И, наконец, Птичьи Признаки большинства последних родственных групп современных птиц необходимо рассматривать в правильном свете. Необходимо обратить внимание, что существует существенная изменчивость в Птичьих Признаках выбранных пяти недавних птиц. В колонке “Все Признаки” Таблицы 4, Птичьи Признаки Ichthyornis и Apsaravis всего на 8 единиц ниже, чем Anas , но соответствующее расстояние от Anas до Crax также составляет 8 единиц. Следовательно, предполагаемое эволюционное изменение, от Ichthyornis до Apsaravis и до современных птиц, очень маленькое маленькое.
Выводы
Трудно избежать вывода, что как потомственная линия от теропод к птице, так и линия от Археоптерикса к современным птицам, являются искусственными. Обе “прогрессии” похожи на пестрые группы, составленные из несвязанных между собой организмов и собранных вместе в последовательность. В конце концов, возвратные признаки либо общие, либо преобладающие, и очень сильно недооценены вследствие огромного количества недостающих данных , невозвратные признаки обычно сами показывают серии резких скачков.
Более того, наиболее птицеподобная часть тероподовой последовательности принадлежит не к предполагаемым предкам первой известной птицы Археоптериксу , но к овираптозавру (oviraptorosaurs ), потомственной ветви “второстепенных нелетающих теропод”. И конечно, статус “второстепенной безполетности” этих овираптозавров (oviraptorosaurs ) подымает вопрос о необходимости в летающих предках. Необходимость для эволюционистов вовлекать этот сложный сценарий событий служит reductio ad absurdum (сведение к абсурду) для эволюционной теории. Вместо того чтобы вовлекать возвратно-поступательный эволюционный процесс из наземных животных к птицам и (в случае с oviraptorosaurs ) обратно к наземным животным, неужели не разумнее просто отказаться от всей эволюции и принять вместо этого специальное сотворение! По причине того, что Создателю не был обязан использовать гнездовую иерархию сотворённых живых существ, по меньшей мере, в каждом случае, не трудно понять, почему эволюционисты сталкиваются с проблемами, пытаясь втиснуть “нептичьи” и “птичьи” признаки в какой-нибудь вид эволюционной линии. Вся ирония данной ситуации заключается в том, что, вопреки предсказаниям тех, кто с удовольствием использует аргумент о стратоморфических промежуточных звеньях, известные тероподы появляются в неправильном месте стратиграфической колонки для того, чтобы играть роль предков птиц.
Современных птиц также полна разрывов в последовательности и возвратных изменениях в признаках. Вцелом, современные птицы не показывают впечатляющей ступенчатой связи с предположительно ранними птицами и еще меньше с Археоптериксом . Изменчивость среди современных птиц значительна, и увеличение диапазона этой изменчивости в несколько раз было бы достаточным для того, чтобы охватить весь ряд Птичьих Признаков, обнаруженных среди птиц и перечисленных в Таблице 2. Это не трудно понять, если учесть тот факт, что дошедшая до наших дней (пережившая Потоп) биосфера, обедненная по сравнению с биосферой, которая существовала до Потопа.
Джон Вудморапп имеет степень Магистра в области геологии и степень Бакалавра биологии от Государственного Среднезападного Университета США. По профессии он преподаватель естественных наук.
Вопросы происхождения жизни и ее развития с древних времен озадачивали ученых. Люди всегда стремились приблизиться к этим тайнам, сделав мир, таким образом, более понятным и предсказуемым. Многие века господствовала точка зрения о божественном начале Вселенной и жизни. Теория эволюции же завоевала почетное место главной и наиболее вероятной версии развития всего живого на нашей планете относительно недавно. Основные положения ее сформулировал Чарльз Дарвин в середине XIX века. Последовавшее за этим столетие подарило миру массу открытий в области генетики и биологии, которые сделали возможным доказать справедливость учения Дарвина, расширить его, объединить с новыми данными. Так появилась синтетическая теория эволюции. Она вобрала в себя все идеи знаменитого исследователя и результаты научных изысканий в различных областях от генетики до экологии.
От особи до класса
Биологическая эволюция представляет собой историческое развитие организмов, основанное на уникальных процессах функционирования генетической информации в определенных условиях окружающей среды.
Начальный этап всех преобразований, в итоге приводящий к появлению нового вида, — это микроэволюция. Такие изменения с течением времени накапливаются и заканчиваются формированием нового более высокого уровня организации живых существ: рода, семейства, класса. Образование надвидовых структур принято называть макроэволюцией.
Аналогичные процессы
Оба уровня в основе своей протекают одинаково. Движущими силами и микро-, и макроизменений являются естественный отбор, изоляция, наследственность, изменчивость. Существенное различие двух процессов в том, что между разными видами практически исключено скрещивание. В результате в основе макроэволюции лежит межвидовой отбор. Огромный вклад в микроэволюцию вносит свободный обмен генетической информацией между особями одного вида.
Схождение и расхождение признаков
Главные направления эволюции могут протекать в нескольких формах. Мощный источник разнообразия жизни — это дивергенция признаков. Она действует как внутри конкретного вида, так и на более высоких уровнях организации. Условия среды и естественный отбор приводят к разделению одной группы на две и более, отличающиеся определенными признаками. На уровне вида дивергенция может быть обратима. В этом случае образовавшиеся популяции вновь сливаются в одну. На более высоких же уровнях процесс необратим.

Еще одна форма — филетическая эволюция, предполагающая преобразование вида без выделения внутри него отдельных популяций. Каждая новая группа является потомком предыдущей и предком для последующей.

Значительный вклад в разнообразие жизни вносит и конвергенция или «схождение» признаков. В процессе развития неродственных групп организмов, находящихся под влиянием одинаковых условий среды, у особей формируются аналогичные органы. Они имеют сходное строение, но разное происхождение и выполняют практически одинаковые функции.
К конвергенции очень близок параллелизм - форма эволюции, когда первоначально дивергирующие группы развиваются похожим образом под влиянием одинаковых условий. Конвергенцию и параллелизм разделяет довольно тонкая грань, и зачастую бывает непросто отнести эволюцию конкретной группы организмов к той или иной форме.
Биологический прогресс
Главные направления эволюции впервые были изложены в трудах А.Н. Северцова. Он предложил выделить понятие биологического прогресса. В работах ученого изложены способы его достижения, а также основные пути и направления эволюции. Идеи Северцова развивал И.И. Шмальгаузен.
Главные направления эволюции органического мира, выделенные учеными, — это биологический прогресс, регресс и стабилизация. По названиям легко понять, чем эти процессы отличаются друг от друга. Прогресс приводит к формированию новых признаков, повышающих степень адаптации организма к среде. Регресс выражается в сокращении численности группы и ее разнообразия, приводя в итоге к вымиранию. Стабилизация влечет закрепление приобретенных признаков и передачи их от поколения к поколению в относительно неизменных условиях.
В более узком смысле, обозначая главные направления органической эволюции, подразумевают именно биологический прогресс и его формы.
Выделяют три основных способа достижения биологического прогресса:
- арогенез;
- аллогенез;
- катагенез.
Арогенез
Данный процесс делает возможным повышение общего уровня организации в результате формирования ароморфоза. Предлагаем выяснить, что понимается под этим понятием. Так, ароморфоз — направление эволюции, приводящее к качественному изменению живых организмов, сопровождающемуся их усложнением и повышением адаптивных свойств. В результате изменения строения становится более интенсивным функционирование особей, они получают возможность использовать новые, ранее незадействованные ресурсы. Как следствие организмы становятся в некотором смысле свободными от условий среды. На более высоком уровне организации их приспособления носят в значительной степени универсальный характер, дающий способность развиваться независимо от окружающих условий.
Хорошим является преобразование кровеносной системы позвоночных: появление четырех камер в сердце и разделение двух кругов кровообращения - большого и малого. Эволюция растений характеризуется значительным скачком вперед в результате формирования пыльцевой трубки и семени. Ароморфозы приводят к появлению новых таксономических единиц: классов, отделов, типов и царств.
Ароморфоз, по Северцову, представляет собой относительно редкое эволюционное явление. Он знаменует собой который, в свою очередь, инициирует прогресс общебиологический, сопровождающийся значительным расширением адаптивной зоны.
Социальный ароморфоз
Рассматривая направления эволюции человеческого рода, некоторые ученые вводят понятие «социальный ароморфоз». Им обозначаются универсальные изменения в развитии социальных организмов и их систем, приводящие к усложнению, большей приспособленности и увеличению взаимовлияния обществ. В число подобных ароморфозов входит, например, появление государства, книгопечатания и компьютерных технологий.
Аллогенез
В ходе биологического прогресса формируются и изменения менее глобального характера. Они составляют суть аллогенеза. У этого направления эволюции (таблица приведена ниже) есть существенное отличие от ароморфоза. Оно не приводит к повышению уровня организации. Главное следствие аллогенеза — это идиоадаптация. По сути, она представляет собой частные изменения, благодаря которым организм получают возможность приспособиться к определенным условиям. эволюции органического мира позволяет близким видам жить в очень разных географических зонах.
Выразительный пример подобного процесса — семейство волчьих. Его виды встречаются в самых разных климатических зонах. Каждый обладает определенным набором приспособлений к своей среде обитания, при этом не превосходя существенно любой другой вид по уровню организации.
Ученые выделяют несколько типов идиоадаптаций:
- по форме (например, обтекаемое тело водоплавающих);
- по окраске (сюда относится мимикрия, предупредительная и ;
- по размножению;
- по передвижению (перепонки водоплавающих, воздушный мешок птиц);
- приспособление к условиям среды.

Различия ароморфоза и идиоадаптации
Некоторые ученые не соглашаются с Северцовым и не видят достаточных причин для различения идиоадаптаций и ароморфозов. Они считают, что степень прогресса может быть оценена только спустя значительное время после появления изменения. По факту сложно осознать, к каким эволюционным процессам приведет новое качество или развитая способность.
Последователи Северцова склоняются к мысли, что под идиоадаптацией следует понимать преобразование формы тела, избыточное развитие или редукцию органов. Ароморфозы же представляют собой существенные изменения в эмбриональном развитии и образование новых структур.
Катагенез
Главные направления эволюции взаимосвязаны и в ходе исторического развития постоянно сменяют друг друга. После кардинальных преобразований в виде ароморфоза или дегенерации наступает период, когда новая группа организмов начинает расслаиваться в результате освоения отдельными ее частями разных географических зон. Начинается эволюция путем идиоадаптаций. Спустя время накопившиеся изменения приводят к новому качественному скачку.
Направление эволюции растений
Современная флора появилась не сразу. Как и все организмы, она прошла длительный путь становления. Эволюция растений включала приобретение нескольких важных ароморфозов. Первым из них было появление фотосинтеза, позволившего примитивным организмам использовать энергию солнечного света. Постепенно в результате преобразований морфологии и фотосинтетических свойств возникли водоросли.
Следующим этапом было освоение суши. Для успешного прохождения «миссии» понадобился еще один ароморфоз — дифференциация тканей. Появились мхи, споровые растения. Дальнейшее усложнение организации связано с преобразованием процесса и способов размножения. Такие ароморфозы, как семязачаток, пыльцевые зерна и, наконец, семя, характеризуют эволюционно более развитые, чем споровые.
Далее пути и направления эволюции растений двигались в сторону еще большей адаптации их к условиям среды, повышению устойчивости к неблагоприятным факторам. В результате появления пестика и зародышевого листка сформировались цветковые или покрытосеменные растения, находящиеся сегодня в состоянии биологического прогресса.

Царство животных
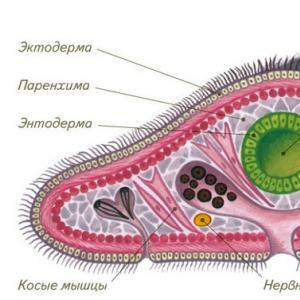
Эволюция эукариотов содержит оформленное ядро) с гетеротрофным типом питания (гетеротрофы не способны создавать органику с помощью хемо- или фотосинтеза) также на первых этапах сопровождалась дифференциацией тканей. Кишечнополостные обладают одним из первых значительных ароморфозов в эволюции животных: у зародышей формируются два слоя, экто- и эндодерма. У круглых и структура уже усложняется. Для них характерен третий зародышевый листок, мезодерма. Этот ароморфоз делает возможным дальнейшую дифференциацию тканей и появление органов.
Следующий этап — формирование вторичной полости тела и дальнейшее разделение его на отделы. уже имеют параподии (примитивный тип конечностей), а также кровеносную и дыхательную системы. Преобразование параподий в членистые конечности и некоторые другие изменения стало причиной появления типа Членистоногих. Уже после их выхода на сушу стали активно развиваться насекомые за счет появления зародышевых оболочек. Они сегодня более всего приспособлены к жизни на земле.
Такие крупные ароморфозы, как формирование хорды, нервной трубки, брюшной аорты и сердца, сделали возможным появление типа Хордовых. Благодаря ряду прогрессивных изменений разнообразие живых организмов пополнилось рыбами, амниотами и рептилиями. Последние из-за наличия зародышевых оболочек перестали зависеть от воды и вышли на сушу.
Далее эволюция идет по пути преобразования кровеносной системы. Возникают теплокровные животные. Адаптации к полету сделали возможным появление птиц. Такие ароморфозы, как четырехкамерное сердце и исчезновение правой дуги аорты, увеличение полушарий переднего мозга и развитие коры, формирование шерстяного покрова и молочных желез и еще ряд изменений привели к появлению млекопитающих. Среди них в процессе эволюции выделились плацентарные животные, и сегодня пребывающие в состоянии биологического прогресса.
Направления эволюции человеческого рода
Вопрос о происхождении и эволюции предков современных людей изучен пока не досконально. Благодаря открытиям палеонтологии и сравнительной генетики изменились уже сложившиеся представления о нашей «родословной». Еще 15 лет назад главенствовала точка зрения, что эволюция гоминид шла по линейному типу, то есть состояла из последовательно сменявших друг друга все более развитых форм: австралопитек, человек умелый, архантроп, неандерталец (палеоантроп), неоантроп (современный человек). Основные направления эволюции человека, как и в случае с другими организмами, приводили к формированию новых адаптаций, повышению уровня организации.

Данные, полученные в последние 10-15 лет, однако, внесли серьезные коррективы в уже сложившуюся картину. Новые находки и уточненная датировка указывают на то, что эволюция имела более сложный характер. Подсемейство Гоминины (относится к семейству Гоминиды) оказалось состоящим из практически вдвое большего числа видов, нежели считалось ранее. Эволюция его не носила линейный характер, а содержала несколько одновременно развивавшихся линий или ветвей, прогрессивных и тупиковых. В разное время вместе сосуществовали три-четыре или больше видов. Сужение этого разнообразия произошло из-за вытеснения эволюционно более развитыми группами других, менее развитых. Например, сейчас уже общепризнанно, что неандертальцы и люди современного типа жили одновременно. Первые не были нашими предками, но представляли собой параллельную ветвь, которую вытеснили более развитые представители гомининов.
Прогрессивные изменения
Несомненными остаются основные ароморфозы, приведшие к процветанию подсемейства. Это прямохождение и увеличение головного мозга. О причинах формирования первого ученые расходятся во мнениях. Долгое время считалось, что это была вынужденная мера, необходимая для освоения открытых пространств. Однако последние данные позволяют утверждать, что предки людей ходили на двух ногах еще в период жизни на деревьях. Такая способность появилась у них сразу после отделения от линии шимпанзе. По одной из версий гоминины изначально передвигались подобно современным орангутанам, стоя двумя ногами на одной ветке и держась руками за другую.
Рост мозга проходил в несколько этапов. Впервые он начался у (человека умелого), который научился изготавливать простейшие орудия. Рост объема головного мозга совпал с увеличением доли мяса в рационе гомининов. Хабилисы, по-видимому, были падальщиками. Следующее увеличение мозга также сопровождалось возрастанием количества мясной пищи и расселением наших предков за пределы родного африканского континента. Ученые предполагают, что рост доли мяса в рационе связан с необходимостью восполнять затраты энергии, уходящей на поддержание работы увеличившегося головного мозга. Предположительно, следующий этап этого процесса совпал с освоением огня: приготовленная пища отличается не только качеством, но и калорийностью, кроме того, существенно уменьшается время, необходимое для пережевывания.

Главные направления эволюции органического мира, действуя на протяжении многих веков, сформировали современную флору и фауну. Движение процесса в сторону адаптации к меняющимся условиям среды привело к огромному разнообразию форм жизни. Главные направления эволюции действуют одинаково на всех уровнях организации, о чем свидетельствуют данные биологии, экологии и генетики.
Класс ПтицыВ мировой фауне обитает около 8600 видов птиц. Это приспособленная к полету веть высших позвоночных, тело которых покрыто перьями. а передние конечности превратились в крылья. Птицы имеют многие черты, сходные с пресмыкающимися: почти полное отсутствие кожных желез; наличие роговых чешуй; сходное строение ряда отделов скелета и мочеполовой системы; тождественное развитие зародыша; артериальную кровь в правой дуге аорты и др.
К чертам, отличающим птиц от пресмыкающихся, можно отнести: прогрессивное развитие органов зрения, слуха и координации движений вместе с соответствующими центрами головного мозга; интенсификацию дыхания, кровообращения и пищеварения совместно с развитием теплоизолирующего покрова и механизмов терморегуляции, обеспечившими постоянную высокую температуру тела; четырехкамерное сердце и полное разделение кругов кровообращения за счет атрофии левой дуги аорты; более совершенные формы размножения; сложный комплекс приспособлений к полету в воздухе, способствующий широким возможностям расселения и добывания пищи. К приспособлениям к полету у птиц можно отнести:
Обтекаемую форму тела с видоизмененными конечностями;
Преобразование передних конечностей в крылья (с этим связано образование киля на грудине, двуногое хождение и появление сложного крестца);
Развитие сложно дифференцированного перьевого покрова;
Наличие облегченных костей;
Развитие системы воздушных мешков и двойного дыхания;
Редукция зубов, замененных роговым клювом, обособление мускульного желудка.
Тело птиц обтекаемой формы, состоит из головы, шеи туловища и конечностей. Шея птиц разной длины, характеризуется большой подвижностью. Небольшая голова заканчивается клювом, состоящим из надклювья и подклювья. Передние конечности преобразованы в крылья. Задние конечности служат опорой всего тела при передвижении по земле. В нижней части они покрыты роговыми чешуями и заканчиваются у большинства птиц четырьмя пальцами.
Покровы представлены тонкой, сухой кожей, лишенной желез. Только над хвостом у большинства птиц расположена особая копчиковая железа. Ее секрет служит для смазывания перьев и защищает их от воды. Снаружи тело птиц покрыто контурными перьями, состоящими из полого стержня, к которому прикреплены опахала. Нижняя часть стержня, погруженная в кожу, называется очином. Опахало состоит из многочисленных длинных бородок первого порядка, на которых находятся бородки второго порядка снабженные мелкими крючочками. Последние соединяют эти бородки между собой. Контурные перья, образующие основную часть несущей плоскости крыла, называют маховыми, образующие плоскость хвоста – рулевые. Под контурными у многих птиц находятся пуховые перья, тонкие стержни которых лишены бородок второго порядка и не образуют сомкнутого опахала. Роль перьевого покрова в жизни птиц разнообразна: перья образуют несущую поверхность крыльев и хвоста, придают обтекаемую форму телу, обеспечивают сохранение тепла и механическую защиту.
Скелет птиц легкий и прочный. В костях имеются полости, заполненные воздухом. Многие кости срастаются между собой – кости черепа, таза. Позвоночник делится на пять отделов: шейный, грудной, поясничный, крестцовый и хвостовой. Шейный отдел обладает значительной подвижностью, остальные же части позвоночника малоподвижны и прочно соединены между собой. Грудные позвонки имеют по паре ребер, сочлененных с грудиной. Ребра имеют крючковидные отростки, которые налегают концами на соседние ребра; это придает грудной клетке прочность. У летающих птиц и пингвинов на грудине расположен киль, к нему прикрепляются сильные грудные мышцы. Задние грудные, поясничные, крестцовые и часть хвостовых позвонков срастаются с костями таза, образуя сложный крестец, создающий прочную опору для ног. Последние хвостовые позвонки срастаются в копчиковую кость, служащую опорой для рулевых перьев.
Череп птиц характеризуется легкостью, имеет большие глазницы и заканчивается клювом. Кости черепа срастаются до полного исчезновения швов между ними.
Плечевой пояс состоит из трех пар костей: вороньих, лопаток, ключиц. Ключицы срастаются своими нижними концами и образуют вилочку, характерную для птиц. Скелет крыла состоит из большой плечевой кости, двух костей предплечья (локтевой и лучевой), сросшихся костей запястья, пястья и редуцированных фаланг трех пальцев. Тазовый пояс состоит из трех пар костей: повздошных, седалищных и лобковых, срастающихся друг с другом. Нижние концы лобковых и седалищных костей не соединяются, поэтому тазовый пояс снизу остается открытым. Это имеет значение для яйцекладки. Скелет задней конечности состоит из бедренной кости, двух сросшихся большой и малой берцовых костей и стопы. В скелет стопы входят цевка (сросшиеся кости плюсны и предплюсны) и фаланги пальцев.
Мускулатура птиц более дифференцирована, чем у пресмыкающихся. Наиболее развиты грудные мышцы, приводящие в движение крылья, а также мышцы задних конечностей, несущие большую нагрузку при хождении.
Пищеварительная система начинается клювом, частично заменяющим зубы; в ротовой полости находится язык. Ротовая полость переходит в короткую глотку, а та в длинный пищевод, имеющий у многих птиц объемистое расширение – зоб. В зобе пища накапливается и размягчается. Нижняя часть пищевода продолжается в железистый желудок, имеющий форму трубки. Там размягченная пища подвергается действию пищеварительных ферментов, выделяемых слизистой оболочкой желудка. Из железистого желудка пища поступает в мышечный желудок, имеющий толстые мускульные стенки и выстланный изнутри роговой кутикулой. Здесь происходит перетирание пищи с помощью и сокращения мышц, и мелких камешков, заглатываемых птицами. Из желудка пища поступает в двенадцатиперстную кишку, куда открываются протоки печени и поджелудочной железы. Тонкая кишка переходит в короткую прямую, открывающуюся в клоаку.
Органы дыхания состоят из легких, воздушных мешков и дыхательных путей, которые начинаются ноздрями, расположенными у основания клюва. Из носовой полости воздух попадает в гортань, а затем в трахею. В месте разделения трахеи на два бронха расположена нижняя, или певчая гортань, характерная для птиц. Она играет роль голосового аппарата. Легкие птиц представляют собой плотные губчатые тела. Бронхи, входящие в легкие, разветвляются. Некоторые бронхи проходят сквозь легкие и оканчиваются тонкостенными воздушными мешками. Они находятся между внутренними органами, между мышцами, под кожей и в полостях трубчатых костей. Воздушные мешки играют большую роль в дыхании, во время полета облегчают массу птицы. При подъеме крыльев (вдох) воздух поступает в легкие, частично отдает кислород и проходит в воздушные мешки. В воздушных мешках окисления крови не происходит. При опускании крыльев (выдох) воздух из мешков снова поступает в легкие. Кровь окисляется в легких как при вдохе, так и при выдохе. Такое дыхание получило название двойного.
Кровеносная система обеспечивает высокий уровень окислительных процессов в теле птиц. В отличие от рептилий кровообращение птиц характеризуется полным разделением артериальной и венозной крови. Четырехкамерное сердце птиц имеет правое и левое предсердия и правый и левый желудочек. Кроме того, исчезает одна из дуг аорты, что исключает возможность смешения крови. Большой круг кровообращения начинается правой дугой аорты, которая огибает сердце и переходит в спинную аорту, идущую вдоль позвоночника. От нее отходят артерии ко всем частям тела. Венозная кровь из передней части тела собирается в парные полые вены, а из задней части тела – в непарную заднюю полую вену. Эти вены впадают в правое предсердие, откуда кровь поступает в правый желудочек.
Малый круг кровообращения начинается легочной артерией, выходящей из правого желудочка. Она несет венозную кровь в легкие, где та окисляется. Из легких артериальная кровь возвращается в левое предсердие по легочным венам. Высокий обмен веществ птиц обеспечивается энергичной работой сердца и быстрой циркуляцией крови по сосудам. Так, у мелких птиц при полете сердце сокращается более 1000 раз/мин.
Органы выделения птиц представлены парными вторичными (тазовыми) почками. Большие по размеру они лежат в углублении тазовых костей. От почек отходят мочеточники, открывающиеся в клоаку. Мочевого пузыря нет, поэтому моча в организме не задерживается.
Нервная система у птиц сложнее, чем у пресмыкающихся. Головной мозг отличается большими размерами полушарий, значительным развитием зрительных бугров и крупным мозжечком. Увеличение среднего мозга связано с интенсификацией зрения у птиц. Большой мозжечок обусловлен полетом, требующим точно координированных движений. От головного мозга отходит 12 пар черепно-мозговых нервов.
Из органов чувств особенно хорошо развито зрение: глаза крупные, снабжены верхним и нижним веками и мигательной перепонкой (третьим веком). Зрение цветное. Большая острота зрения обеспечивается двойной аккомодацией: изменением формы хрусталика и расстояния между хрусталиком и сетчаткой. Органы слуха состоят из внутреннего и среднего уха, барабанная перепонка несколько углублена. Органы обоняния развиты слабо.
Размножение происходит путем откладывания яиц. Птицы раздельнополы; половой диморфизм выражен; оплодотворение внутреннее. Половая система самца состоит из двух семенников и семяпроводов, открывающихся в клоаку. У самое развиваются левый яичник и яйцевод, который открывается в клоаку. После оплодотворения яйцо медленно движется по яйцеводу и покрывается белковой оболочкой. В нижней, расширенной, части яйцевода (матке) оно покрывается тонкими подкорлуповыми оболочками и скорлупой и через узкое влагалище попадает в клоаку. Развитие зародыша идет под защитой зародышевых оболочек.
Развитие зародыша птиц начинается из небольшого зародышевого диска, находящегося на поверхности желтка. Яйцо птиц имеет крупные размеры, так как содержит много питательных веществ в виде желтка и белка. Желток держится на канатиках (состоят из густого белка) в центре яйца. Благодаря небольшому удельному весу зародышевый диск при любом положении яйца располагается сверху. Это имеет большое значение при насиживании, так как зародыш получает больше теплоты. Белок покрыт двумя подскорлуповыми оболочками, расходящимися на тупом конце яйца и образующими воздушную камеру. Снаружи яйцо покрыто известковой скорлупой, имеющей большое количество пор. Через поры происходит газообмен между зародышем и внешней средой. Скорлупа покрыта тончайшей надскорлуповой оболочкой, защищающей яйцо от проникновения бактерий. Развитие зародыша идет под защитой зародышевых оболочек.
Насиживание у птиц продолжается различное время: у голубя 16-18 суток, у курицы – 21 сутки, у африканского страуса – до 40 суток. После насиживания из яиц вылупляются птенцы. У одних птиц птенцы покрыты густым пухом и способны к самостоятельному образу жизни (выводковые птицы). У других – птенцы не покрыты перьями и долгое время выкармливаются родителями (птенцовые птицы). К выводковым относят курообразных, гусеобразных, журавлей, страусов. К птенцовым – воробьиных, дятлообразных, голубей. Существует промежуточная группа – это чайки, дневные хищники; по развитию их можно отнести к выводковым, а по характеру питания – к птенцовым.
Оседлые птицы живут на определенных территориях, кочующие – совершают перелеты к югу, перелетные – улетают в теплые страны и проводят там зиму.
К классу Птицы принадлежат надотряды Пингвины, Бескилевые, или Страусовые, Килегрудые. Географическое распространение птиц широко. И они приспособились к разнообразным условиям существования.
Основную массу птиц культурных ландшафтов (парков, садов, лугов, полей) составляют представители отряда воробьиных. Это насекомоядные птицы мелких и средних размеров. К ним относятся воробьи, синицы, ласточки, соловьи, сороки, галки, жаворонки и др.
Птицы, обитающие в лесах, приспособились лазать по деревьям, когти у них очень острые и сильно загнуты. Могут питаться личинками насекомых, лесными ягодами. К птицам леса относят дятлов, глухарей, тетеревов и др.
Некоторые птицы приспособились к обитанию в водоемах, болотах: на ногах у них имеются плавательные перепонки; кроме того, существует копчиковая железа, выделения которой нужны для защиты от воды. К водоплавающим относят утку-крякву, пингвинов и др.
На открытых пространствах степей и пустынь обитают птицы, передвигающиеся главным образом бегом или шагом: крылья короткие и широкие, ноги сильные с короткими пальцами. К ним относятся журавли, страусы, дрофы. Питаются растительной пищей, насекомыми, иногда мелкими грызунами.
Среди хищников различают дневных, охотящихся днем (грифы, ястребы, соколы, орлы и др.), и ночных (совы, филины). Питаются они животными, иногда рыбой, насекомыми. Имеют острые когти, мощный клюв и хорошее зрение.
Птицы регулируют численность насекомых, мелких грызунов. Участвуют в распространении семян, опылении. Многие украшают природные ландшафты и оживляют их своем пением. Польза и вред птиц относительны. Они поддерживают природные очаги заболеваний человека как хранители и прокормители переносчиков этих заболеваний; птицы принимают участие в циркуляции вирусных заболеваний (орнитозов, гриппа, энцефалитов).
 |
|
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
Зачет
Тип
:
тест по теме «Пресмыкающиеся и Птицы»
Предмет
:
биология
Класс
:
7
Составитель
заданий
:
Гарейшина Инна Георгиевна
Среди крупных видов крокодилов отдельные особи могут доживать до 120 лет, кошки редко живут дольше 12 лет. Во сколько раз дольше может жить крокодил? Ответ записать числом.
Пингвины – обитатели южного полушария Земли. Самый крупный пингвин – императорский, рост около 1 м, приплывает гнездиться на самый холодный континент – Антарктиду. Самый мелкий – малый пингвин, ростом всего около 40 см, гнездится практически на экваторе, на Галапагосских островах. На сколько сантиметров императорский пингвин выше малого? Ответ записать в сантиметрах, числом.
Самая большая птица на Земле – африканский страус. Его рост (более 2,5 м) превышает рост человека. Летать он не может. Обладая мощными ногами, эта птица способна бежать со скоростью 60 км в час. Сколько километров страус пробежит за 10 минут? Ответ записать числом.
Масса куриного яйца примерно 60 граммов, а яйца африканского страуса – в 25 раз больше. Какова масса страусиного яйца? Ответ записать числом.
В классе пресмыкающихся или рептилий наибольшее число видов в отряде ящериц, примерно 3500 видов, а в отряде змей на 500 видов меньше. Отряд черепах, примерно, в 10 раз малочисленнее, чем змей, а в отряде крокодилов в 15 раз меньше видов, чем у черепах. К отряду клювоголовых относятся гаттерии, сохранившиеся только в Новой Зеландии, их в 10 раз меньше, чем крокодилов. Сколько видов содержит каждый из отрядов рептилий? Ответом будет являться четыре числа.
Мухоловка-пеструшка приносит птенцам около 500 беспозвоночных животных в день. Сколько беспозвоночных она скормит за 16 дней своим птенцам, пока они не вылетят из гнезда? Ответ записать числом.
У серой цапли в полной кладке 3-7 зеленовато-голубых яиц. Сколько птенцов может появиться у пары цапель за три года, если в каждой кладке будет по 6 яиц? Ответ записать числом.
Воробьев можно считать типичными зерноядными птицами, однако своих птенцов они выкармливают беспозвоночными животными, в основном насекомыми. В период выкармливания птенцов, всего за две недели, пара воробьев скармливает своим прожорливым детишкам около 1,4 кг насекомых, пауков и моллюсков. Сколько граммов беспозвоночных воробьи скармливают птенцам за 1 день? Ответ записать числом.
Сокращение численности галапагосских черепах началось с XVIII века, в настоящее время популяции восстанавливаются человеком. Самая большая сухопутная черепаха живет на Галапагосских островах, длина ее панциря может достигать полутора метров, а вес до двухсот килограмм. Самая большая морская черепаха на 50 см длиннее, а ее вес больше, чем у сухопутного вида, в 2,5 раза. Какова масса и длина тела морской черепахи? Ответом будет являться два числа.
ПРИЗНАК А) четырехкамерное сердце Б) кожа сухая, тонкая, покрыта роговыми чешуями и пластинами В) хорошо развита забота о потомстве
Г) кровь в сердце смешанная
Д) температура тела высокая и постоянная
Е) трехкамерное сердце с неполной перегородкой в желудочке
1)Пресмыкающиеся 2) Птицы
Ему хвост помогает охотиться. Удар его хвоста опасен даже для крупных животных. Покрытый роговыми пластинками, он тяжелый, как бревно. Мощным хвостом это животное сбивает жертву с ног, подбрасывает вверх и ловит огромной зубастой пастью. 1) игуана, 2) варан, 3) крокодил, 4) анаконда, 5) молох. Ответом будет являться один номер.
Приспособлениями птиц к полету можно считать:
Видоизмененные конечности
Хорошее обоняние
Один круг кровообращения и легочное дыхание
Полые кости в скелете
Наличие двенадцатиперстной и прямой кишок
Отсутствие мочевого пузыря
Выберите правильные утверждения. Ответом будет являться ряд чисел.
Дыхательная поверхность легких ящерицы больше, чем у тритона
У всех пресмыкающихся трехкамерное сердце
Пресмыкающиеся откладывают яйца
У пресмыкающихся северных районов чаще встречается живорождение
В сердце ящерицы имеется полная межжелудочковая перегородка
У пресмыкающихся в коже много потовых и сальных желез
В пищевой цепи коршун является вторичным потребителем, потому, что он:
Гетеротроф
Хищник
Использует энергию солнца
Регулирует численность поедаемых им животных
Минерализует органические остатки
Питается слабыми и больными животными
Существенными эволюционными, прогрессивными приобретениями птиц являются:
Головной мозг из пяти отделов
Интенсивный обмен веществ
Смешанная кровь в сердце
Костный череп
Клюв без зубов
Легкость и прочность скелета
Правильные ответы
| Номер задания | Ответ |
| 1 | 10 |
| 2 | 60 |
| 3 | 10 |
| 4 | 1500 |
| 5 | 3000, 300, 20, 2 |
| 6 | 8000 |
| 7 | 18 |
| 8 | 100 |
| 9 | 200, 500 |
| 10 | 2,1,2,1,2,1 |
| 11 | 3 |
| 12 | 1,4,6 |
| 13 | 1,3,4 |
| 14 | 2,4,6 |
| 15 | 2,5,6 |